
林业科学 ›› 2023, Vol. 59 ›› Issue (8): 60-73.doi: 10.11707/j.1001-7488.LYKX20220709
何苏诚,宋军阳*( )
)
收稿日期:2022-10-20
出版日期:2023-08-25
发布日期:2023-10-16
通讯作者:
宋军阳
E-mail:281168046@qq.com
基金资助:
Sucheng He,Junyang Song*()
Received:2022-10-20
Online:2023-08-25
Published:2023-10-16
Contact:
Junyang Song
E-mail:281168046@qq.com
摘要:
目的: 血皮槭种子存在深度生理胚休眠,制约其种群恢复和生产应用。探究血皮槭种子休眠机制,寻求解除休眠的方法,可为血皮槭及其他植物的种子休眠机制研究和生产实践提供参考。方法: 对血皮槭种子自然状态下萌发过程进行动态观测,研究其自然休眠时长;通过3~12个月低温层积、切离胚以及不同浓度赤霉素溶液处理,探寻打破血皮槭种子休眠的方法;利用休眠的血皮槭种子各部分浸提液处理小麦种子做发芽试验,分析各处理组小麦种子的发芽势、发芽率和发芽指数,分析血皮槭种子中抑制其萌发的物质存在部位;通过代谢组技术,探究影响血皮槭种子休眠的具体物质。结果: 1)自然状态下,血皮槭种子休眠时间约为29个月;血皮槭种子经3~12个月的低温层积、切离胚和200、400、800 mg·L–1浓度的赤霉素溶液处理均不能打破休眠。2)休眠血皮槭种子的种翅和外种皮浸提液处理的小麦种子发芽势和发芽率高于对照组;休眠血皮槭种子种胚浸提液处理的小麦种子发芽势、发芽率和发芽指数显著低于对照组(P<0.01);打破休眠的血皮槭种子种胚浸提液处理的小麦种子发芽势、发芽率和发芽指数显著高于休眠的种子种胚浸提液处理组。3)血皮槭休眠的种子种胚和打破休眠的种子种胚中显著差异代谢物分别为289个和215个,通过二级谱图匹配这些显著差异代谢物在正离子模式下有146个,负离子模式下有109个;差异倍数(|log2FC|)最大的20个代谢物中,有19个在种子打破休眠后含量显著减少,它们主要分布在脂肪酰基类和苯及其取代衍生物类中;通过一级谱图,定性了一类显著差异代谢物——酚磷酸;脂肪酰基类和酚类代谢物在影响血皮槭种子休眠中起关键作用,抑制血皮槭种子发芽的物质是脂肪酰基类的2-羟基辛酸、茉莉酸、反式-2-辛烯酸和十三碳二元酸,以及酚类的甲基生姜酚、黄尿酸、香草酸甲酯。结论: 血皮槭种子休眠属于深度生理胚休眠,休眠时间约为29个月;血皮槭种子的种翅、外种皮和内种皮对其萌发没有抑制作用,且种翅和外种皮中含有促进种子发芽的物质;抑制血皮槭种子发芽的物质存在于种胚中;血皮槭种子休眠并不是单一的内源性物质造成的,可能是多种物质的互作或者叠加的结果。
中图分类号:
何苏诚,宋军阳. 血皮槭种子休眠特性及休眠原因[J]. 林业科学, 2023, 59(8): 60-73.
Sucheng He,Junyang Song. Characteristics and Mechanism of Seed Dormancy of Acer griseum[J]. Scientia Silvae Sinicae, 2023, 59(8): 60-73.
表1
低温层积处理、切离胚处理和不同浓度的赤霉素溶液处理对血皮槭种子打破休眠的影响①"
| 低温层积 Low temperature stratification | 切离胚 Embryo sectioning | 赤霉素浓度 Concentrations of GA3/(mg·L–1) | 发芽率 Germination rate(%) |
| 否No | 否No | 0 | 0 |
| 否No | 否No | 200 | 0 |
| 否No | 否No | 400 | 0 |
| 否No | 否No | 800 | 0 |
| 否No | 是Yes | 0 | 0 |
| 否No | 是Yes | 200 | 0 |
| 否No | 是Yes | 400 | 0 |
| 否No | 是Yes | 800 | 0 |
| 是Yes | 否No | 0 | 0 |
| 是Yes | 是Yes | 0 | 0 |

图1
血皮槭种子4个不同部分浸提液对小麦种子萌发的抑制作用比较 (A)小麦种子第3天发芽情况;(B)小麦种子第7天发芽情况。A1:种翅浸提液;A2:外种皮浸提液;A3:内种皮浸提液;A4:休眠种胚浸提液;UP:超纯水。"


图2
血皮槭休眠的种子种胚和打破休眠的种子种胚浸提液对小麦种子萌发的抑制作用比较 (A)小麦种子第3天发芽情况;(B)小麦种子第7天发芽情况。A4:休眠种胚浸提液; A5:打破休眠种胚浸提液;UP:超纯水。"


图3
血皮槭休眠种子各部分浸提液和打破休眠种子种胚浸提液对小麦种子萌发的影响 (A)发芽势,(B)发芽率,(C)发芽指数。A1:种翅浸提液; A2:外种皮浸提液; A3:内种皮浸提液; A4:休眠种胚浸提液; A5:打破休眠种子种胚浸提液; UP:超纯水。不同小写字母表示处理之间在P<0.01差异极显著。"
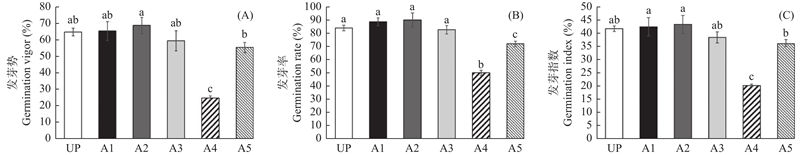

图4
质量控制主成分分析 (A)正离子模式下质量控制PCA,(B)负离子模式下质量控制PCA。CK:打破休眠的种子种胚;T:休眠的种子种胚;QC:所有样本混合。PC1坐标表示第一主成分,括号中的百分比表示第一主成分对样品差异的贡献值;PC2坐标表示第二主成分,括号中的百分比表示第二主成分对样品差异的贡献值。图中有颜色的点分别表示各个样品,同一组样本距离越近,说明样本重复性越好。QC样本越密集分布,说明数据越可靠。"
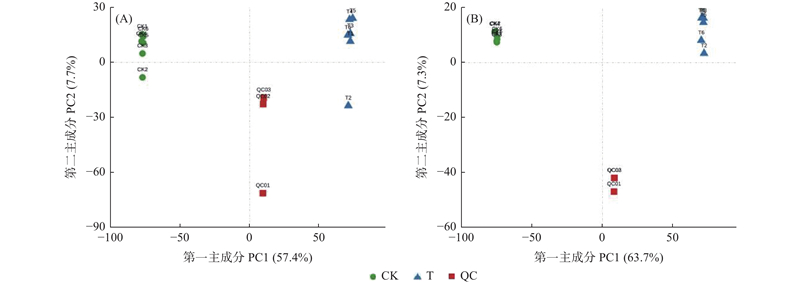

图5
样本相关性热图 (A)正离子模式下样本相关性热图;(B)负离子模式下样本相关性热图。图中每一行和每一列都代表样本,每个格内的数值表示对应的两个样本的Pearson相关系数,数值越接近1,颜色越深,表示两个样本相关性越强。"
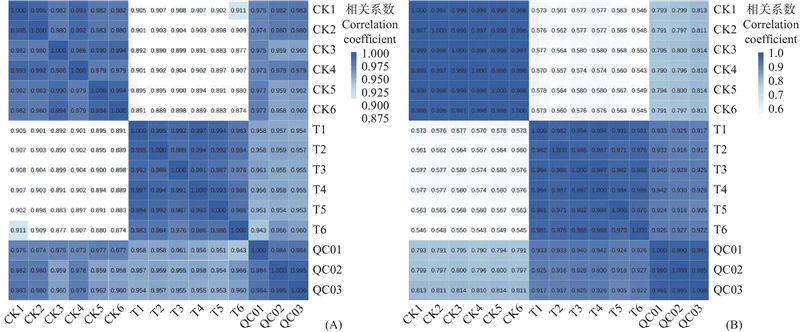

图6
OPLS-DA得分 to1: 正交成分得分值Orthogonal component score value. t1: 主要成分得分值Major component score value. RMSEE:均方根误差Root mean square error. ort:正交组分数目Number of orthogonal components. Pre:主成分+正交主成分Principal component +Orthogonal principal component. R2X(cum):模型(对X变量数据集)可解释度Explainability of the model (for X variable data set). R2Y(cum):模型(对Y变量数据集)可解释度Explainability of the model (for Y variable data set). Q2(cum):模型可预测度Model predictability."

图7
KEGG富集圈 第1圈:富集前20的通路(pathway),圈外为差异代谢物数目的坐标尺;不同的颜色代表不同的通路A级分类(A class)。第2圈:差异代谢物背景中该通路的数目以及Q值;差异代谢物背景数量越多条形越长,Q值越小颜色越红。第3圈:上下调差异代谢物比例条形图,深紫色代表上调差异代谢物比例,浅紫色代表下调差异代谢物比例;下方显示具体的数值。第4圈:各通路的富集因子(rich factor)值(该通路中差异代谢物数量除以该通路中所有数量),背景网格线,每一格代表0.1。"


图8
差异倍数(|log2FC|)Top20的代谢物聚类热图 图中每一行代表一种代谢物,每一列代表一个样本,将20个代谢物利用R包pheatmap(v1.0.12)(Kolde et al., 2018)进行数据z-score标准化后进行聚类分析并画热图,可直观展示代谢物在样本中的表达规律,颜色越红,表示代谢物丰度越高,颜色越蓝代表代谢物丰度越低。根据代谢物丰度分别对代谢物和样本进行聚类,同一分支下的样本或代谢物表达模式更相近。"
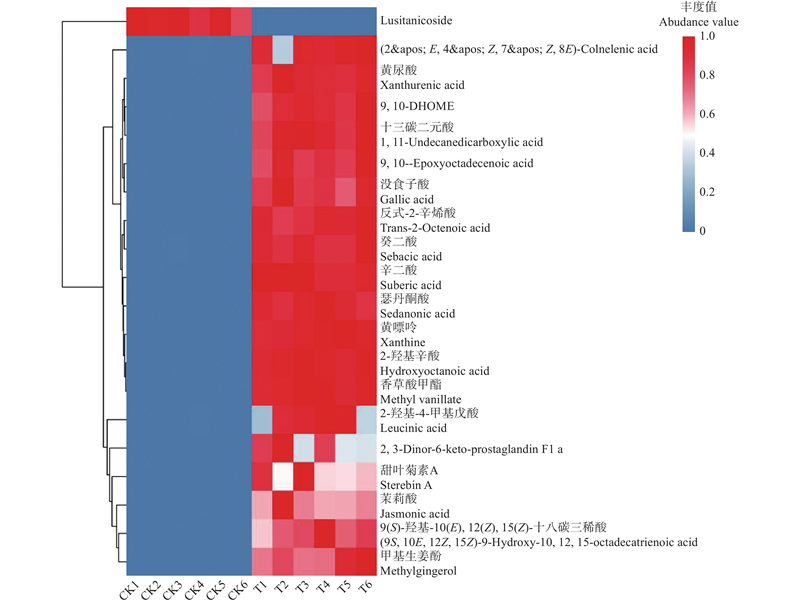
表2
差异倍数(|log2FC|)Top20的显著差异代谢物①"
| 二级谱图匹配的 差异代谢物 Differences metabolites(MS2) | 化学物质登录号 CAS | 代谢物的分类 CLASS | 对照组(CK)积分定量值均值 Control_mean | 处理组(T)积分定量值均值 Case_mean | 差异倍数取log2 log2FC(case_mean/control_mean) |
| 2-羟基辛酸 Hydroxyoctanoic acid | 617-73-2 | 脂肪酰基 Fatty acyls | 1 | 116 323 933.50± 3 275 143.49 | 26.79 |
| 甲基生姜酚 Methylgingerol | — | 苯及其取代衍生物 Benzene and substituted derivatives | 1 | 51 566 708.15± 7 972 029.56 | 25.62 |
| 黄尿酸 Xanthurenic acid | 59-00-7 | 喹啉及其衍生物 Quinolines and derivatives | 1 | 33 927 291.88± 1 936 667.29 | 25.02 |
| 反式-2-辛烯酸 trans-2-Octenoic acid | 1871-67-6 | 脂肪酰基Fatty acyls | 1 | 25 985 461.12± 1 572 720.82 | 24.63 |
| 茉莉酸 Jasmonic acid | 59366-47-1 | 脂肪酰基 Fatty acyls | 1 | 24 092 770.25± 5 100 433.30 | 24.52 |
| 十三碳二元酸 1,11-Undecanedicarboxylic acid | 505-52-2 | 脂肪酰基 Fatty acyls | 1 | 18 552 823.64± 1 485 075.26 | 24.15 |
| 香草酸甲酯 Methyl vanillate | 3943-74-6 | 苯及其取代衍生物 Benzene and substituted derivatives | 1 | 17 917 489.67± 659 781.08 | 24.09 |
| 甜叶菊素A Sterebin A | 107647-14-3 | 孕烯醇酮脂类 Prenol lipids | 29 979.24± 20 623.82 | 28 021 234.01± 8 625 740.76 | 9.87 |
| 9,10-DHOME | — | 脂肪酰基 Fatty acyls | 273 046.59± 60 964.19 | 203 012 632.00± 15 799 278.61 | 9.54 |
| 没食子酸 Gallic acid | 149-91-7 | 苯及其取代衍生物 Benzene and substituted derivatives | 366 515.84± 124 845.54 | 234 320 907.50± 22 477 504.64 | 9.32 |
| 黄嘌呤 Xanthine | 69-89-6 | 咪唑并嘧啶类 Imidazopyrimidines | 200 441.72± 64 282.21 | 84 786 371.81± 2 852 164.17 | 8.72 |
| (2'E,4'Z,7'Z,8E)-Colnelenic acid | — | — | 120 076.71± 153 245.34 | 36 596 004.06± 10 850 047.06 | 8.25 |
| Lusitanicoside | 499-35-4 | 含氧有机物 Organooxygen compounds | 16 050 537.74± 1 297 721.17 | 68 719.84± 18 060.10 | ?7.87 |
| 辛二酸 Suberic acid | 505-48-6 | 脂肪酰基 Fatty acyls | 182 053.34± 83 626.64 | 39 659 265.40± 1 759 173.88 | 7.77 |
| 癸二酸 Sebacic acid | 111-20-6 | 脂肪酰基 Fatty acyls | 112 787.29± 63 607.65 | 19 396 356.98± 963 237.47 | 7.43 |
| 2,3-Dinor-6-keto-prostaglandin F1 a | — | 脂肪酰基 Fatty acyls | 302 167.62± 55 097.29 | 48 728 392.67± 19 852 430.85 | 7.33 |
| 2-羟基-4-甲基戊酸 Leucinic acid | 498-36-2 | 脂肪酰基 Fatty acyls | 856 742.28± 479 120.63 | 125 609 896.80± 55 570 190.11 | 7.20 |
| 9(S)-羟基-10(E),12(Z),15(Z)-十八碳三稀酸 (9S,10E,12Z,15Z)-9-Hydroxy-10,12,15-octadecatrienoic acid | — | — | 294 080.13± 47 110.04 | 31 460 665.89± 5 505 953.97 | 6.74 |
| 9,10-Epoxyoctadecenoic acid | — | 脂肪酰基 Fatty acyls | 6 817 811.33± 1 794 368.54 | 683 861 405.70± 61 296 093.44 | 6.65 |
| 瑟丹酮酸 Sedanonic acid | 6697-07-0 | 生酮酸及其衍生物 Keto acids and derivatives | 323 831.19± 169 883.62 | 31 587 321.59± 2 416 888.82 | 6.61 |

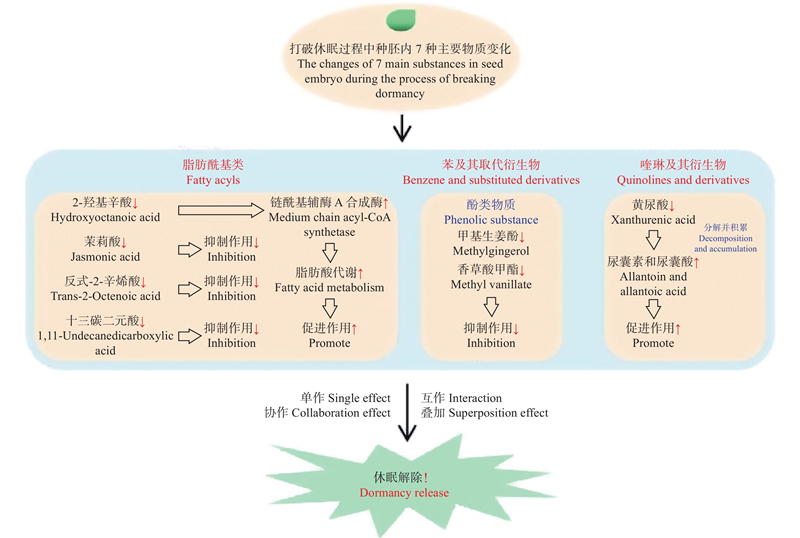
图9
血皮槭种子打破休眠的模式 红色箭头向下:下调;红色箭头向上:上调。 Red arrow down: down-regulated; Red arrow up: up-regulated."
| 曹 山, 蒋璐瑶, 李丽红, 等. 毛果杨中链酰基辅酶A合成酶的克隆及酶学分析. 北京林业大学学报, 2016, 38 (7): 9- 15. | |
| Cao S, Jiang L Y, Li L H, et al. Cloning and enzymatic analysis of medium-chain acyl coenzyme A synthetase in Populus trichocarpa . Journal of Beijing Forestry University, 2016, 38 (7): 9- 15. | |
| 陈 丽, 李 冰, 王 梅, 等. 血皮槭的愈伤组织培养研究. 北方园艺, 2010, (8): 170- 172. | |
| Chen L, Li B, Wang M, et al. Study on callus culture of Acer griseum . Northern Horticulture, 2010, (8): 170- 172. | |
| 陈 朋, 于雪丹, 张川红, 等. 中国特有种血皮槭的天然更新. 林业科学, 2013, 49 (3): 159- 164. | |
| Chen P, Yu X D, Zhang C H, et al. Natural regeneration of Acer griseum, an endemic species in China . Scientia Silvae Sinicae, 2013, 49 (3): 159- 164. | |
| 杜丽雁, 张春涛, 高 强, 等. 血皮槭播种育苗技术. 林业实用技术, 2005, (3): 24. | |
| Du L Y, Zhang C T, Gao Q, et al. Sowing and cultivation technique of Acer griseum . Forest Science and Technology, 2005, (3): 24. | |
| 付其迪. 2020. 血皮槭叶绿体基因组变异及与近缘种系统进化分析. 北京: 中国林业科学研究院. | |
| Fu Q D. 2020. Variation of chloroplast genomes in Acer griseum and its phylogenetic relationship with related species. Beijing: Chinese Academy of Forestry.[in Chinese] | |
| 郭幸飞, 乔 琦, 李 婷. 特有珍稀植物血皮槭种子的生物学特征和贮藏特性研究. 种子, 2017, 36 (12): 20- 24. | |
| Guo X F, Qiao Q, Li T. The study on the seed biology and seed storage characteristics of Acer griseum, an endemic and rare plant in China . Seed, 2017, 36 (12): 20- 24. | |
| 孙佳伟, 郑勇奇, 于雪丹, 等. 濒危植物血皮槭花部特征和交配系统. 林业科学, 2022, 58 (6): 47- 55. | |
| Sun J W, Zheng Y Q, Yu X D, et al. Floral traits and mating system of endangered species Acer griseum . Scientia Silvae Sinicae, 2022, 58 (6): 47- 55. | |
| 孙 圣. 2014. 基于SSR标记的血皮槭天然群体遗传变异研究. 北京: 中国林业科学研究院. | |
| Sun S. 2014. Study on genetic variation of natural populations in Acer griseum based on SSR markers. Beijing: Chinese Academy of Forestry.[in Chinese] | |
| 王佳慧. 2015. 特有濒危种血皮槭天然群体生长状况及分子谱系地理初步研究. 北京: 中国林业科学研究院. | |
| Wang J H. 2015. Preliminary study on growth performance and molecular phylogeography of natural populations in an endangered maple, Acer griseum (Dicotyledoneae: Sapindaceae), endemic to China. Beijing: Chinese Academy of Forestry.[in Chinese] | |
| 汪 松, 解 焱. 2004. 中国物种红色名录. 第一卷, 红色名录. 北京: 高等教育出版社. | |
| Wang S, Xie Y. 2004. China species red list.Vol.1 Beijing: Higher Education Press.[in Chinese] | |
| 叶学敏. 2017. 濒危植物血皮槭种群动态预测及潜在适生区模拟研究. 北京: 中国林业科学研究院. | |
| Ye X M. 2017. Prediction of population dynamics and simulation of potentially suitable distribution in endangered species Acer griseum. Beijing: Chinese Academy of Forestry.[in Chinese] | |
| 张川红, 王佳慧, 于雪丹, 等. 特有种野生血皮槭的生长状况. 林业科学研究, 2015, 28 (3): 421- 425. | |
| Zhang C H, Wang J H, Yu X D, et al. Growth performance of wild paperbark maple (Acer griseum) . Forest Research, 2015, 28 (3): 421- 425. | |
| 张川红, 郑勇奇, 吴 见, 等. 血皮槭种子休眠机制研究. 植物研究, 2012, 32 (5): 573- 577. | |
| Zhang C H, Zheng Y Q, Wu J, et al. Mechanisms of seed dormancy of Acer griseum . Bulletin of Botanical Research, 2012, 32 (5): 573- 577. | |
| 中国科学院中国植物志编辑委员会. 1981. 中国植物志, 第46卷. 北京: 科学出版社. | |
| Flora Reipublicae Popularis Sinicae Editorial Committee of Chinese Academy of Sciences. 1981. Flora Reipublicae Popularis Sinicae, 46. Beijing: Science Press.[in Chinese] | |
| Alamillo J M, Díaz-Leal J L, Sánchez-Moran M V, et al. 2010. Molecular analysis of ureide accumulation under drought stress in Phaseolus vulgaris L. Plant, Cell & Environment, 33(11): 1828−1837. | |
| Baskin J M, Baskin C C. 1998. Seeds–ecology, biogeography, and evolution of dormancy and germination. San Diego, CA, USA: Academic Press. | |
|
Baskin J M, Baskin C C. A classification system for seed dormancy. Seed Science Research, 2004, 14 (1): 1- 16.
doi: 10.1079/SSR2003150 |
|
|
Berestetzky V, Dathe W, Daletskaya T, et al. Jasmonic acid in seed dormancy of Acer tataricum . Biochemie Und Physiologie Der Pflanzen, 1991, 187 (1): 13- 19.
doi: 10.1016/S0015-3796(11)80178-2 |
|
|
Bylesjo M, Rantalainen M, Cloarec O, et al. OPLS discriminant analysis: combining the strengths of PLS-DA and SIMCA classification. Journal of Chemometrics, 2006, 20, 341- 351.
doi: 10.1002/cem.1006 |
|
| Chang Y W, Zhao C X, Zhu Z, et al. Metabolic profiling based on LC/MS to evaluate unintended effects of transgenic rice with cry1Ac and sck genes . Plant Molecular Biology, 2012, 78 (4/5): 477- 487. | |
|
Chase M W, Christenhusz M J M, Fay M F, et al. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Botanical Journal of the Linnean Society, 2016, 181 (1): 1- 20.
doi: 10.1111/boj.12385 |
|
| Corbineau F, Lecat S, Côme D. Dormancy of three cultivars of oat seeds (Avena sativa L.) . Seed Science & Technology, 1986, 14, 725- 735. | |
|
Corbineau F, Rudnicki R M, CoMe D. The effects of methyl jasmonate on sunflower (Helianthus annuus L.) seed germination and seedling development . Plant Growth Regulation, 1988, 7 (3): 157- 169.
doi: 10.1007/BF00028238 |
|
|
Dunn W B, Broadhurst D, Begley P, et al. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nature Protocols, 2011, 6 (7): 1060- 1083.
doi: 10.1038/nprot.2011.335 |
|
|
Fernández-Valverde M, Reglero A, Martinez-Blanco H, et al. Purification of Pseudomonas putida acyl coenzyme A ligase active with a range of aliphatic and aromatic substrates . Applied and Environmental Microbiology, 1993, 59(4), 1149- 1154.
doi: 10.1128/aem.59.4.1149-1154.1993 |
|
|
Finch-Savage W E, Leubner-Metzger G. Seed dormancy and the control of germination. New Phytologist, 2006, 171(3), 501- 523.
doi: 10.1111/j.1469-8137.2006.01787.x |
|
|
Finkelstein R, Reeves W, Ariizumi T, et al. Molecular aspects of seed dormancy. Annual Review of Plant Biology, 2008, 59, 387- 415.
doi: 10.1146/annurev.arplant.59.032607.092740 |
|
|
Fujihara S, Yamaguchi M. Effects of allopurinol [4-hydroxypyrazolo (3, 4-d) pyrimidine] on the metabolism of allantoin in soybean plants. Plant Physiology, 1978, 62(1), 134- 138.
doi: 10.1104/pp.62.1.134 |
|
|
Fulda M, Shockey J, Werber M, et al. Two long-chain acyl-CoA synthetases from Arabidopsis thaliana involved in peroxisomal fatty acid beta-oxidation . Plant Journal, 2002, 32(1), 93- 103.
doi: 10.1046/j.1365-313X.2002.01405.x |
|
| Gao F, Ayele B T. Functional genomics of seed dormancy in wheat: advances and prospects. Frontiers in Plant Science, 2014, 5, 458. | |
|
Gleiser G, Picher M C, Veintimilla P, et al. Seed dormancy in relation to seed storage behaviour in Acer . Botanical Journal of the Linnean Society, 2004, 145(2), 203- 208.
doi: 10.1111/j.1095-8339.2003.00276.x |
|
|
Graeber K, Nakabayashi K, Miatton E, et al. Molecular mechanisms of seed dormancy. Plant, Cell & Environment, 2012, 35(10), 1769- 1786.
doi: 10.1111/j.1365-3040.2012.02542.x |
|
|
Holdsworth M J, Bentsink L, Soppe W J J. Molecular networks regulating Arabidopsis seed maturation, after-ripening, dormancy and germination. New Phytologist, 2008, 179(1), 33- 54.
doi: 10.1111/j.1469-8137.2008.02437.x |
|
|
Ishimata N, Ito H, Tai A. Structure-activity relationships of vanillic acid ester analogs in inhibitory effect of antigen-mediated degranulation in rat basophilic leukemia RBL-2H3 cells. Bioorganic & Medicinal Chemistry Letters, 2016, 26(15), 3533- 3536.
doi: 10.1016/j.bmcl.2016.06.028 |
|
|
Kanehisa M, Goto S. KEGG: kyoto encyclopedia of genes and genomes. Nucleic Acids Research, 2000, 28(1), 27- 30.
doi: 10.1093/nar/28.1.27 |
|
|
Kasuya F, Igarashi K, Fukui M. Inhibition of a medium chain acyl-CoA synthetase involved in glycine conjugation by carboxylic acids. Biochemical Pharmacology, 1996, 52(10), 1643- 1646.
doi: 10.1016/S0006-2952(96)00563-1 |
|
| Kolde R, Kolde M R. 2018. Package ‘pheatmap’. R Package, 1. | |
|
Kong L, Wang F, Si J, et al. Water-soluble phenolic compounds in the coat control germination and peroxidase reactivation in Triticum aestivum seeds . Plant Growth Regulation, 2008, 56(3), 275- 283.
doi: 10.1007/s10725-008-9307-2 |
|
|
Kuras M. Cytochemical localization of phenolic compounds in columella cells of the root cap in seeds of Brassica napus—changes in the localization of phenolic compounds during germination . Annals of Botany, 1999, 84(2), 135- 143.
doi: 10.1006/anbo.1999.0884 |
|
| Lenoir C, Corbineau F, Côme D. Barley (Hordeum vulgare) seed dormancy as related to glumella characteristics . Physiologia Plantarum, 2010, 68(2), 301- 307. | |
|
Lisec J, Schauer N, Kopka J, et al. Gas chromatography mass spectrometry-based metabolite profiling in plants. Nature Protocols, 2006, 1(1), 387- 396.
doi: 10.1038/nprot.2006.59 |
|
|
Mothes K. The metabolism of urea and ureides. Canadian Journal of Botany, 1961, 39 (7): 1785- 1807.
doi: 10.1139/b61-158 |
|
|
Naseri B, Tabari M, Phartyal S S, et al. Deep physiological dormancy in seeds of Balkan maple (Acer hyrcanum): a rare tree in the Hyrcanian Mountain forests of Iran . Seed Science and Technology, 2018, 46(3), 473- 482.
doi: 10.15258/sst.2018.46.3.04 |
|
|
Nelson S K, Ariizumi T, Steber C M. Biology in the dry seed: transcriptome changes associated with dry seed dormancy and dormancy loss in the Arabidopsis GA-insensitive sleepy1-2 mutant . Frontiers in Plant Science, 2017, 8, 2158.
doi: 10.3389/fpls.2017.02158 |
|
|
Nojavan-Asghari M, Ishizawa K. Inhibitory effects of methyl jasmonate on the germination and ethylene production in cocklebur seeds. Journal of Plant Growth Regulation, 1998, 17(1), 13- 18.
doi: 10.1007/PL00007006 |
|
| Nonogaki H. Seed dormancy and germination-emerging mechanisms and new hypotheses. Frontiers in Plant Science, 2014, 5, 233. | |
|
Nourimand M, Todd C D. There is a direct link between allantoin concentration and cadmium tolerance in Arabidopsis . Plant Physiology and Biochemistry, 2019, 135, 441- 449.
doi: 10.1016/j.plaphy.2018.11.016 |
|
| Oracz K, Karpiński S. Phytohormones signaling pathways and ROS involvement in seed germination. Frontiers in Plant Science, 2016, 7, 864. | |
| Pawłowski T A. Proteome analysis of Norway maple (Acer platanoides L.) seeds dormancy breaking and germination:influence of abscisic and gibberellic acids . BMC Plant Biology, 2009, 9, 48. | |
| Phartyal S S, Thapliyal R C, Nayal J S, et al. 2003. Seed dormancy in Himalayan maple (Acer caesium) II: bioassay of inhibitors. Seed Science and Technology, 31(1): 13−20. | |
|
Pinfield N J, Dungey N O. Seed dormancy in Acer: an assessment of the rôle of the structures covering the embryo . Journal of Plant Physiology, 1985, 120(1), 65- 81.
doi: 10.1016/S0176-1617(85)80123-1 |
|
| Pinfield N J, Stutchbury P A. 1990b. Seed dormancy in Acer: the role of testa-imposed and embryo dormancy in Acer velutinum. Annals of Botany, 66(2): 133−137. | |
|
Pinfield N J, Stutchbury P A, Bazaid S A, et al. Abscisic acid and the regulation of embryo dormancy in the genus Acer . Tree Physiology, 1990a, 6(1), 79- 85.
doi: 10.1093/treephys/6.1.79 |
|
|
Qi M Q, Upadhyaya M K, Furness N H, et al. Mechanism of seed dormancy in Cynoglossum officinale L . Journal of Plant Physiology, 1993, 142(3), 325- 330.
doi: 10.1016/S0176-1617(11)80430-X |
|
| Reinbothe H. 1962. Urea, ureidea, and guanidines in plants. Annual Review Plant Physiology, 13: 129−150. | |
|
Saccenti E, Hoefsloot H C J, Smilde A K, et al. Reflections on univariate and multivariate analysis of metabolomics data. Metabolomics, 2014, 10(3), 361- 374.
doi: 10.1007/s11306-013-0598-6 |
|
|
Sińska I, Gladon R J. Ethylene and the removal of embryonal apple seed dormancy. HortScience, 1984, 19(1), 73- 75.
doi: 10.21273/HORTSCI.19.1.73 |
|
| Steinberg S J, Morgenthaler J, Heinzer A K, et al. Very long-chain acyl-CoA synthetases. Human "bubblegum" represents a new family of proteins capable of activating very long-chain fatty acids. Journal of Biological Chemistry, 2000, 275 (45): 35162- 35169. | |
| Thevenot E A. 2016. ropls: PCA, PLS (-DA) and OPLS (-DA) for multivariate analysis and feature selection of omics data. https://bioconductor.statistik.tu-dortmund.de/packages/3.6/bioc/vignettes/ropls/inst/doc/ropls-vignette.pdf | |
|
Tognacca R S, Servi L, Hernando C E, et al. Alternative splicing regulation during light-induced germination of Arabidopsis thaliana seeds . Frontiers in Plant Science, 2019, 10, 1076.
doi: 10.3389/fpls.2019.01076 |
|
| Vordtriede S A, Vordtriede P B, Schulz K E, et al. 1999. Embryo germination in Acer ginnala Maxim. and the activity of an endogenous exudate. Transactions of the Illinois State Academy of Science, 92: 59−67. | |
| Wang W C, Chen S Y, Zhang X Z. 2017. The complete chloroplast genome of the endangered Chinese paperbark maple, Acer griseum (Sapindaceae). Conservation Genetics Resources, 9: 527–529. | |
| Warnes G R. 2007. gmodels: various R programming tools for model fitting. R package version 2.16.2. https://CRAN.R-project.org/package=gmodels. | |
|
Webb D P, Dumbroff E. Factors influencing the stratification process in seeds of Acer saccharum . Canadian Journal of Botany, 1969, 47(10), 1555- 1563.
doi: 10.1139/b69-222 |
|
| Webb D P, Wareing P F. 1972. Seed dormancy in Acer pseudoplatanus L.: the role of the covering structures. Journal of Experimental Botany, 23(3): 813−829. | |
| Yang N, Guo X, Wu Y, et al. 2018. The inhibited seed germination by ABA and MeJA is associated with the disturbance of reserve utilizations in Astragalus membranaceus. Journal of Plant Interactions, 13: 388−397. | |
|
Yildiz K, Muradoglu F, Yilmaz H. The effect of jasmonic acid on germination of dormant and nondormant pear (Pyrus communis L.) seeds . Seed Science and Technology, 2008, 36(3), 569- 574.
doi: 10.15258/sst.2008.36.3.06 |
|
| Yildiz K, Yazici C, Muradoglu F. Effect of jasmonic acid on germination dormant and nondormant apple seeds. Asian Journal of Chemistry, 2007, 19, 1098- 1102. |
| [1] | 孙佳伟,郑勇奇,于雪丹,夏新合,赵玉贤,武宇霞,张川红. 濒危植物血皮槭花部特征和交配系统[J]. 林业科学, 2022, 58(6): 47-55. |
| [2] | 杨治华,陈雪梅,陈春桦,张丽苗,董智,谭雪,李昌晓. 池杉种皮特性和种子休眠原因[J]. 林业科学, 2022, 58(4): 11-21. |
| [3] | 耿瑜欣,李红姣,郑建伟,张芹,于丽娜,李佳秋,李保会. 皂荚与野皂荚刺中次生代谢物的差异[J]. 林业科学, 2022, 58(4): 82-94. |
| [4] | 陈新,王敏,傅茂润,王贵芳,相昆,刘庆忠,焦文晓,张美勇,许海峰. 核桃炭疽病发生相关的酚类物质代谢分析[J]. 林业科学, 2021, 57(10): 71-80. |
| [5] | 王艳梅,张小雪,朱秀征,张志华,李志,耿晓东,蔡齐飞,刘震. 林木种子休眠与萌发机制研究进展[J]. 林业科学, 2021, 57(10): 128-144. |
| [6] | 尹德洁,布凤琴,徐艳芳,黄彪,李成. 单叶蔓荆种子休眠特性与解除方法[J]. 林业科学, 2020, 56(12): 157-165. |
| [7] | 陆小雨,陈竹,唐菲,傅松玲,任杰. 转录组与代谢组联合解析红花槭叶片中花青素苷变化机制[J]. 林业科学, 2020, 56(1): 38-53. |
| [8] | 刘果, 陈鸿鹏, 吴志华, 彭彦, 谢耀坚. 南美油藤种子发育过程的代谢组学和转录组学联合分析[J]. 林业科学, 2019, 55(5): 169-179. |
| [9] | 候亚会, 严善春, 李志强. 对苯二酚对台湾乳白蚁诱食效果及其诱导代谢的响应[J]. 林业科学, 2018, 54(9): 97-103. |
| [10] | 曹亚兵, 翟晓巧, 邓敏捷, 赵振利, 范国强. 泡桐丛枝病发生与代谢组变化的关系[J]. 林业科学, 2017, 53(6): 85-93. |
| [11] | 陈香波, 刘杨, 赵明水, 涂淑萍. 极度濒危树种羊角槭的种胚发育与休眠解除[J]. 林业科学, 2017, 53(4): 65-73. |
| [12] | 闫芳, 张恩和, 王勤礼, 毛著鸿. 野生黄瑞香种子的休眠机制与破除方法[J]. 林业科学, 2016, 52(4): 30-37. |
| [13] | 陈朋;于雪丹;张川红;郑勇奇;孙圣;程蓓蓓;朱从波. 中国特有种血皮槭的天然更新[J]. 林业科学, 2013, 49(3): 159-164. |
| [14] | 尚旭岚;徐锡增;方升佐. 青钱柳种子休眠机制[J]. 林业科学, 2011, 47(3): 68-74. |
| [15] | 沈海龙 杨玲 张建瑛 冯丹丹 范少辉. 花楸树种子休眠影响因素与萌发特性研究[J]. 林业科学, 2006, 42(10): 133-138. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||