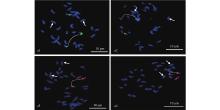
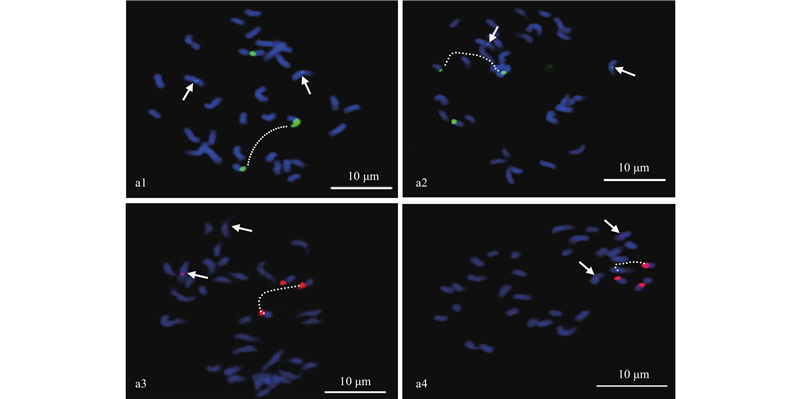
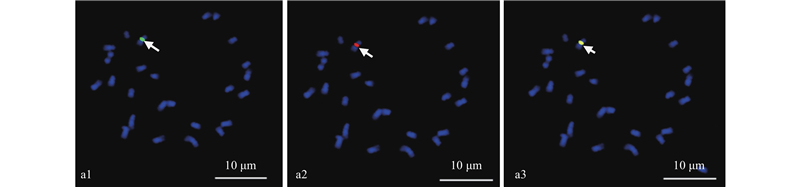
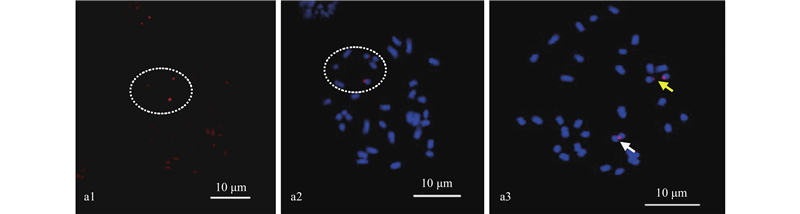
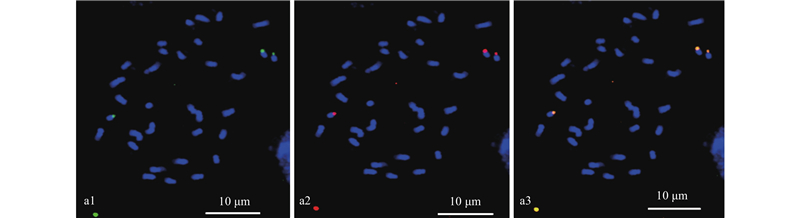
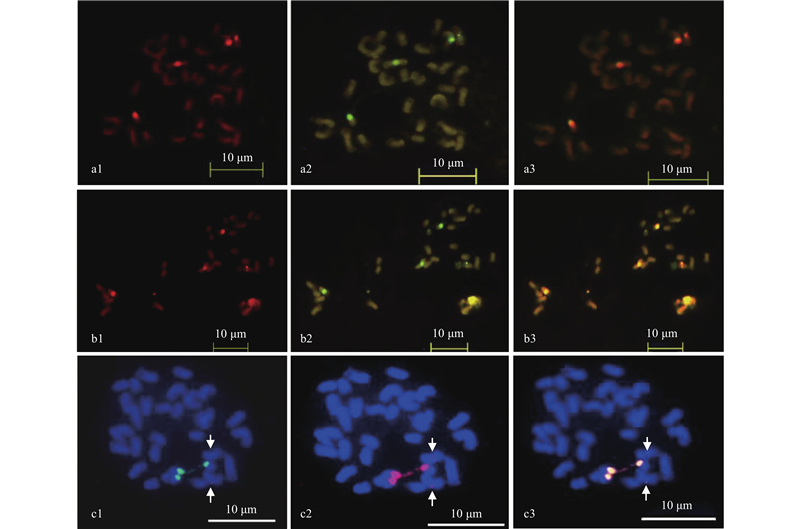
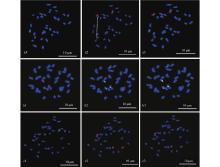
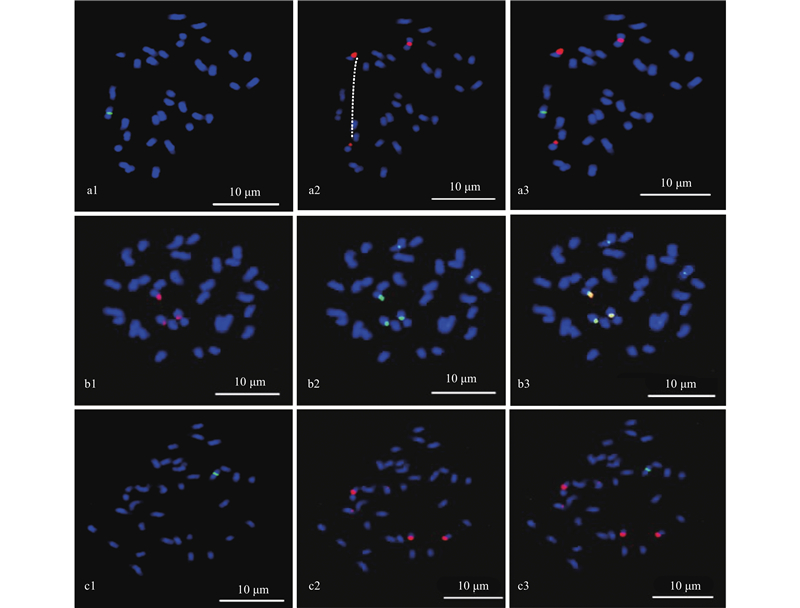
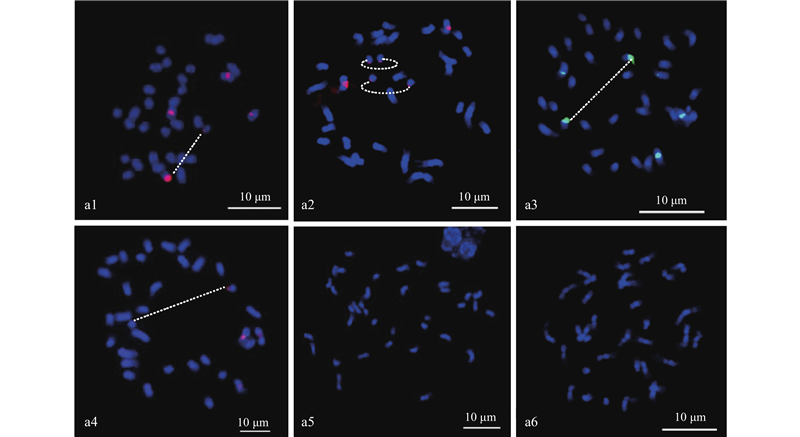
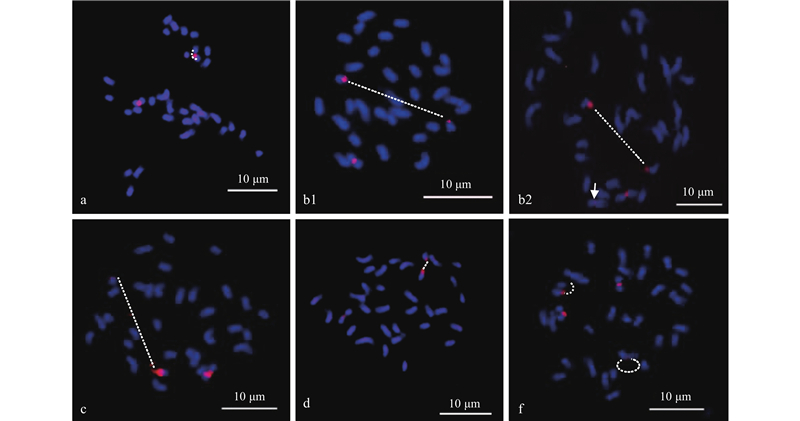
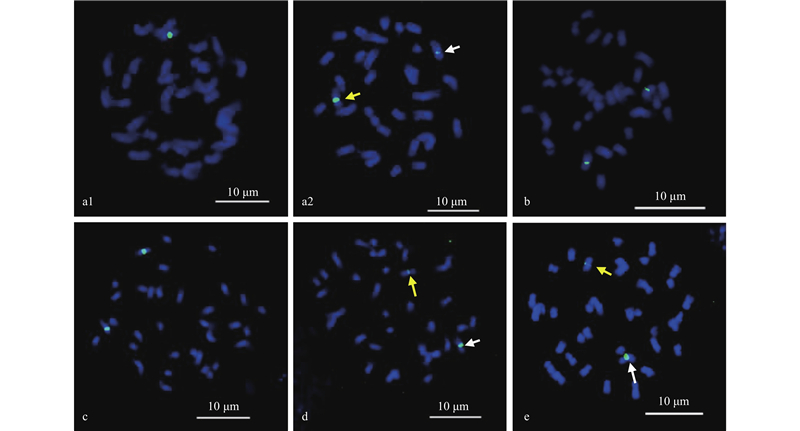
|
陈瑞阳, 李秀兰, 宋文芹, 等. 2003. 中国主要经济植物基因组染色体图谱Ⅳ. 北京: 科学出版社, 1−628.
|
|
Chen R Y, Li X L, Song W Q, et al. 2003. Chromosome atlas of major economic plants genome in China (Version IV): Chromosome atlas of various bamboo species. Beijing: Science Press, 1−628.[in Chinese]
|
|
杜 培. 2017. 小麦、百萨偃麦草和花生染色体荧光原位杂交寡核苷酸探针(套)开发与应用. 南京: 南京农业大学.
|
|
Du P. 2017. Development and application of wheat, thinopyrwn bessarabicwn and peanut oligonucleotide and multiplex probes for fluorescence in situ hybridization. Nanjing: Nanjing Agricultural University.[in Chinese]
|
|
刘茂春, 黎章矩. 中国山核桃属一新种. 浙江林学院学报, 1984, 1 (1): 41- 42.
|
|
Liu M C, Li Z J. A new species of Carya from China . Journal of Zhejiang Forestry College, 1984, 1 (1): 41- 42.
|
|
吕芳德, 杨 帆, 张日清. 山核桃属部分种的核型分析. 中南林学院学报, 2002, 22 (1): 47- 49.
|
|
Lü F D, Yang F, Zhang R Q. Karyotypes of three Carya Nutt. species . Journal of Central South Forestry University, 2002, 22 (1): 47- 49.
|
|
徐川梅. 2017. 10个山核桃种染色体核型及基因组类型分析. 北京: 中国林业科学研究院.
|
|
Xu C M. 2017. Analysis of karyotype and genome type of ten Carya species. Beijing: Chinese Academy of Forestry.[in Chinese]
|
|
Albert P S, Zhang T, Semrau K, et al. Whole-chromosome paints in maize reveal rearrangements, nuclear domains, and chromosomal relationships. Proceedings of the National Academy of Sciences of the United States of America, 2019, 116 (5): 1679- 1685.
doi: 10.1073/pnas.1813957116
|
|
Braz G T, He L, Zhao H N, et al. Comparative oligo-FISH mapping: an efficient and powerful methodology to reveal karyotypic and chromosomal evolution. Genetics, 2018, 208 (2): 513- 523.
doi: 10.1534/genetics.117.300344
|
|
Cai Z X, Liu H J, He Q Y, et al. Differential genome evolution and speciation of Coix lacryma-jobi L . and Coix aquatica Roxb. hybrid guangxi revealed by repetitive sequence analysis and fine karyotyping. BMC Genomics, 2014, 15, 1025.
|
|
Chen R Y, Song W Q, Liang G L. Chromosome atlas of Chinese fruit trees and their close wild relatives. Beijing:Internatinal Academic Publishers, 1993, 1, 351- 353.
|
|
Cheng Z K, Buell C R, Wing R A, et al. Toward a cytological characterization of the rice genome. Genome Research, 2002, 11 (12): 2133- 2141.
|
|
Chung K S, Weber J A, Hipp A L. 2011. Dynamics of chromosome number and genome size variation in a cytogenetically variable sedge (Carex scoparia var. scoparia, Cyperaceae). American Journal of Botany, 98(1): 122−129.
|
|
Dong F, Song J, Naess S K, et al. Development and applications of a set of chromosome-specific cytogenetic DNA markers in potato. Theoretical and Applied Genetics, 2000, 101 (7): 1001- 1007.
doi: 10.1007/s001220051573
|
|
Du P, Cui C H, Liu H, et al. Development of an oligonucleotide dye solution facilitates high throughput and cost-efficient chromosome identification in peanut. Plant Methods, 2019, 15, 69.
doi: 10.1186/s13007-019-0451-7
|
|
Du P, Li L A, Zhang Z X, et al. Chromosome painting of telomeric repeats reveals new evidence for genome evolution in peanut. Journal of Integrative Agriculture, 2016, 15 (11): 2488- 2496.
doi: 10.1016/S2095-3119(16)61423-5
|
|
Du P, Li L N, Liu H, et al. High-resolution chromosome painting with repetitive and single-copy oligonucleotides in Arachis species identifies structural rearrangements and genome differentiation . BMC Plant Biology, 2018, 18 (1): 240.
doi: 10.1186/s12870-018-1468-1
|
|
Du P, Zhuang L F, Wang Y Z, et al. Development of oligonucleotides and multiplex probes for quick and accurate identification of wheat and Thinopyrum bessarabicum chromosomes . Genome, 2017, 60 (2): 93- 103.
doi: 10.1139/gen-2016-0095
|
|
HanY H, Zhang Z H, Thammapichai P, et al. Chromosome-specific painting in Cucumis species using bulked oligonucleotides. Genetics, 2015, 200 (3): 771- 779.
doi: 10.1534/genetics.115.177642
|
|
He Q Y, Cai Z X, Hu T H, et al. Repetitive sequence analysis and karyotyping reveals centromere-associated DNA sequences in radish (Raphanus sativus L.) . BMC Plant Biology, 2015, 15, 105.
doi: 10.1186/s12870-015-0480-y
|
|
Huang Y J, Xiao L H, Zhang Z, et al. The genomes of pecan and Chinese hickory provide insights into Carya evolution and nut nutrition . Gigascience, 2019, 8 (5): 1- 17.
|
|
Jiang J M. Fluorescence in situ hybridization in plants: recent developments and future applications. Chromosome Research, 2019, 27 (3): 153- 165.
doi: 10.1007/s10577-019-09607-z
|
|
Liu X Y, Sun S, Wu Y, et al. Dual-color oligo-FISH can reveal chromosomal variations and evolution in Oryza species . Plant Journal, 2019, 101 (1): 112- 121.
|
|
Manos P S, Stone D E. Evolution, phylogeny, and systematics of the Juglandaceae. Annals of the Missouri Botanical Garden, 2001, 88 (2): 231- 269.
doi: 10.2307/2666226
|
|
Mondin M, Aguiar-Perecin, M L. Heterochromatin patterns and ribosomal DNA loci distribution in diploid and polyploid Crotalaria species (Leguminosae, Papilionoideae) and inferences on karyotype evolution . Genome, 2011, 54 (9): 718- 726.
|
|
Moraes A P, Simões A O, Alayon D I O, et al. Detecting mechanisms of karyotype evolution in Heterotaxis (Orchidaceae) . PLoS One, 2016, 11, e0165960.
doi: 10.1371/journal.pone.0165960
|
|
Roa F, Guerra M. 2012. Distribution of 45S rDNA sites in chromosomes of plants: structural and evolutionary implications. BMC Evolutionary Biology, 12: 225.
|
|
Rocha L C, de Oliveira Bustamante F, Silveira R A, et al. Functional repetitive sequences and fragile sites in chromosomes of Lolium perenne L . Protoplasma, 2015, 252 (2): 451- 60.
doi: 10.1007/s00709-014-0690-4
|
|
Vasconcelos E V, Vasconcelos S, Ribeiro T, et al. Karyotype heterogeneity in Philodendron s. l. (Araceae) revealed by chromosome mapping of rDNA loci . PLoS One, 2018, 13 (11): 1- 15.
|
|
Xin H Y, Zhang T, Wu Y F, et al. An extraordinarily stable karyotype of the woody Populus species revealed by chromosome painting . Plant Journal, 2020, 101 (2): 253- 264.
doi: 10.1111/tpj.14536
|
|
Yakovlev S S, Godelle B, Zoldos V, et al. Evolutionary implications of heterochromatin and rDNA in chromosome number and genome size changes during dysploidy: a case study in Reichardia genus . PLoS One, 2017, 12 (8): e0182318.
doi: 10.1371/journal.pone.0182318
|
|
Yu F, Zhao X W, Chai J, et al. Chromosome-specific painting unveils chromosomal fusions and distinct allopolyploid species in the Saccharum complex . New Phytologist, 2021, 233 (4): 1953- 1965.
|
|
Zhang J B, Li R Q, Xiang X G, et al. Integrated fossil and molecular data reveal the biogeographic diversification of the Eastern Asian-Eastern North American disjunct hickory genus (Carya Nutt.) . PLoS One, 2013, 8 (7): e70449.
doi: 10.1371/journal.pone.0070449
|