
Scientia Silvae Sinicae ›› 2025, Vol. 61 ›› Issue (12): 34-48.doi: 10.11707/j.1001-7488.LYKX20250230
• Frontiers and hot topics • Previous Articles
Long Zhou1,4,Maifang Zhang2,Qiang Dong2,Huinan Zhu1,3,*( ),Jinliang Liu1,4,*()
),Jinliang Liu1,4,*()
Received:2025-04-14
Revised:2025-09-08
Online:2025-12-25
Published:2026-01-08
Contact:
Huinan Zhu,Jinliang Liu
E-mail:geldwxh@163.com;liujinliang2016@nwafu.edu.cn
CLC Number:
Long Zhou,Maifang Zhang,Qiang Dong,Huinan Zhu,Jinliang Liu. Characteristics of Soil Microbial-Derived Carbon Changes in Pinus tabuliformis Forest, Quercus wutaishanica Forest, and Their Mixed Forest[J]. Scientia Silvae Sinicae, 2025, 61(12): 34-48.
Table 1
Various stand types information"
| 林分类型 Stand type | 优势乔木 Dominant tree species | 平均胸径 Mean DBH/ cm | 平均树高 Mean tree height/m | 海拔 Elevation/ m | 坡向 Aspect | 坡度 Slope/ (°) | 郁闭度 Canopy density | 灌木优势种Dominant shrub species | 草本优势种Dominant grass species |
| 油松纯林 Pinus tabuliformis forests | 油松 Pinus tabuliformis | 21.41±4.33 | 12.28±0.28 | 1 202 | 西南206° Southwest 206° | 23 | 0.60 | 野蔷薇Rosa multiflora、白刺花Sophora davidii、忍冬Lonicera japonica | 细秆薹草Carex capillaris、黄精Polygonatum sibiricum |
| 辽东栎纯林 Quercus wutaishanica forests | 辽东栎 Quercus wutaishanica | 21.41±0.34 | 11.41±0.75 | 1 603 | 北4° North 4° | 27 | 0.75 | 忍冬Lonicera japonica、红瑞木Lonicera japonica、荚蒾Lonicera japonica | 细秆薹草Carex capillaris、茜草 Rubia cordifolia |
| 油松-辽东栎 混交林 Pinus tabuliformis- Quercus wutaishanica mixed forests | 油松 Pinus tabuliformis | 25.90±2.70 | 14.59±0.83 | 1 535 | 西258° West 258° | 15 | 0.75 | 灰栒子Lonicera japonica、忍冬Lonicera japonica、红瑞木Lonicera japonica | 细秆薹草Carex capillaris、白茅草Imperata cylindrica |
| 辽东栎 Quercus wutaishanica | 18.90±3.67 | 11.55±1.55 |
Table 2
Soil nutrient status and stoichiometric ratios of different stand types"
| 土壤养分特征 Soil nutrient characteristics | 油松纯林 Pinus tabuliformis forests | 辽东栎纯林 Quercus wutaishanica forests | 油松-辽东栎混交林 P. tabuliformis-Q. wutaishanica mixed forests |
| SOC/ (g·kg?1) | 13.17±0.89a | 16.05±1.67 a | 13.95±0.50 a |
| TN/ (g·kg?1) | 1.00±0.04 a | 1.40±0.10 a | 1.47±0.23 a |
| TP/ (g·kg?1) | 0.53±0.05 a | 0.64±0.02 a | 0.69±0.03 a |
| AP/ (mg·kg?1) | 0.18±0.01b | 0.28±0.03a | 0.22±0.05a |
| NH4+-N/ (mg·kg?1) | 2.32±0.22b | 4.60±0.34a | 5.03±0.18a |
| NO3?-N/ (mg·kg?1) | 0.30±0.05ab | 0.25±0.03b | 0.42±0.05a |
| C∶N | 13.13±0.72a | 11.36±0.60b | 10.27±0.30ab |
| C∶P | 26.12±4.45a | 25.21±2.33a | 20.49±1.37a |
| N∶P | 0.45±0.04a | 0.31±0.02b | 0.29±0.04b |
Table 3
Soil microbial PLFA biomass content in different stand types"
| 微生物活体碳类别 Microbial PLFA biomass | 油松纯林 P. tabuliformisforests | 辽东栎纯林 Q. wutaishanica forests | 油松-辽东栎混交林 P. tabuliformis-Q. wutaishanica mixed forests |
| 放线菌门Actinobacteria/ (mg·kg?1) | 1.34±0.04b | 1.32±0.16b | 1.98±0.30a |
| 厚壁菌门Firmicutes/ (mg·kg?1) | 1.86±0.02b | 2.20±0.29b | 2.95±0.09a |
| 丛枝菌根真菌Arbuscular mycorrhizal fungi/ (mg·kg?1) | 0.24±0.01b | 0.38±0.06ab | 0.47±0.07a |
| 子囊菌和担子菌门Ascomycota and Basidiomycota/ (mg·kg?1) | 0.40±0.06a | 0.42±0.08a | 0.57±0.02a |
| 接合菌门Zygomycota/ (mg·kg?1) | 0.69±0.01b | 0.83±0.12b | 1.31±0.10a |
| 革兰氏阴性菌Gram-negative bacteria/ (mg·kg?1) | 3.44±0.33a | 3.65±0.51a | 5.38±0.85a |
| 革兰氏阳性菌Gram-positive bacteria/ (mg·kg?1) | 3.17±0.12b | 3.57±0.45ab | 4.91±0.70a |
| 细菌Bacteria/ (mg·kg?1) | 6.61±0.42b | 7.18±0.96ab | 10.29±1.54a |
| 真菌Fungi/ (mg·kg?1) | 1.48±0.08b | 1.74±0.23b | 2.58±0.12a |
| 真菌∶细菌Ratio of bacteria to fungi | 0.22±0.01a | 0.26±0.02a | 0.24±0.02a |
| 微生物Microbial/ (mg·kg?1) | 9.73±0.58b | 10.96±1.59ab | 15.09±2.06a |

Fig.1
Microbial PLFA biomass carbon content per unit mass of organic carbon in different stand types"
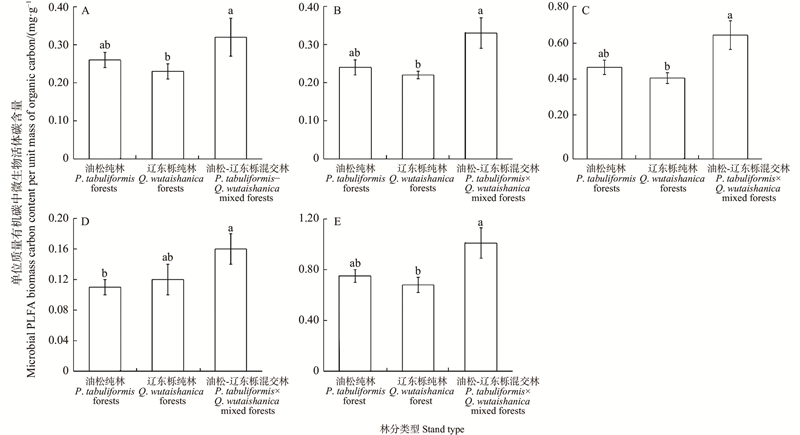
Table 4
Soil microbial necromass carbon and amino sugar contents in different stand types"
| 项目Item | 油松纯林 P. tabuliformis forests | 辽东栎纯林 Q. wutaishanica forests | 油松-辽东栎混交林 P. tabuliformis-Q. wutaishanica mixed forests |
| MurN/ (mg·kg?1) | 19.66±1.04b | 21.25±1.40b | 30.81±4.36a |
| GalN/ (mg·kg?1) | 209.88±13.00a | 298.43±20.27a | 302.59±47.23a |
| GluN/ (mg·kg?1) | 566.37±7.83b | 603.85±52.32ab | 709.70±10.67a |
| BNC/ (mg·kg?1) | 884.66±46.59b | 956.28±62.78b | 1 386.65±196.15a |
| FNC/ (mg·kg?1) | 4 844.90±57.31b | 5 161.81±455.46ab | 6 000.98±80.10a |
| MNC/ (mg·kg?1) | 5 729.49±103.60b | 6 118.09±510.16b | 7 354.68±136.04a |
| F∶BN | 5.53±0.24a | 5.39±0.26a | 4.69±0.68a |

Fig.2
Contribution of soil microbial necromass carbon to soil organic carbon"
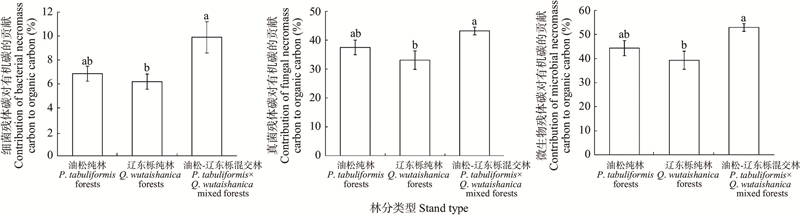
Table 5
Relative abundance of genes encoding differential CAZy enzymes for plant- and microbial-derived components in different stand types %"
| 组分Components | 油松纯林 P. tabuliformis forests | 辽东栎纯林 Q. wutaishanica forests | 油松-辽东栎混交林 P. tabuliformis-Q. wutaishanica mixed forests | |
| 植物源 Plant-derived | 半纤维素Hemicellulose | 48.49±0.43b | 51.93±1.07a | 49.86±0.73ab |
| 纤维素Cellulose | 7.40±0.16b | 8.09±0.09a | 8.01±0.17a | |
| 木质素Lignin | 30.56±0.32a | 27.75±0.86b | 29.44±0.49ab | |
| 总和Total | 86.45±0.09b | 87.76±0.30a | 87.31±0.27ab | |
| 细菌源 Bacteria-derived | 肽聚糖Peptidoglycan | 9.54±0.09a | 8.28±0.39b | 8.59±0.44ab |
| 真菌源 Fungi-derived | 几丁质Chitin | 2.93±0.11a | 2.79±0.05a | 3.02±0.16a |
| 葡聚糖Glucose | 1.08±0.06a | 1.16±0.13a | 1.08±0.07a | |
| 总和Total | 4.01±0.08a | 3.96±0.16a | 4.09±0.21a | |

Fig.3
Cluster analysis of relative abundance of microbial CAZyme genes that degrade plant and microbial sources of carbon"
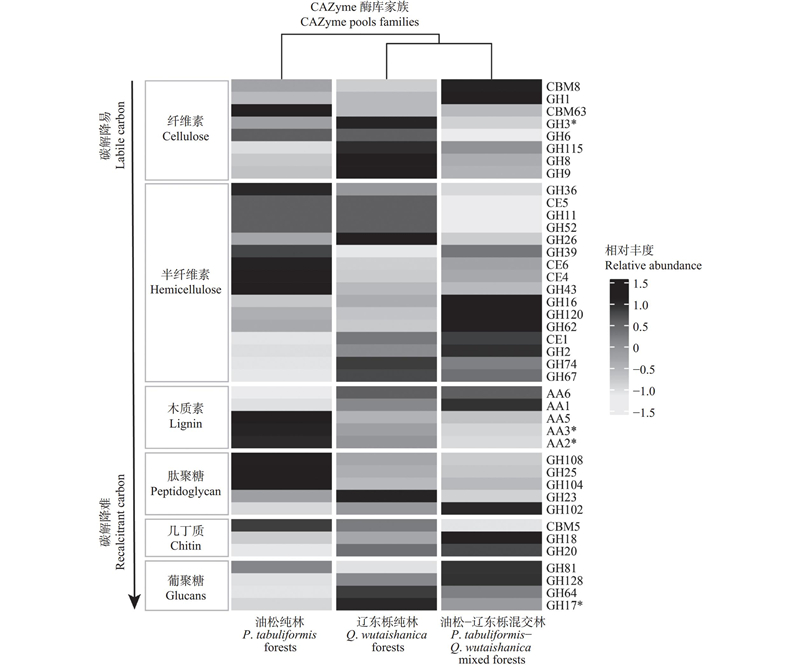
Table 6
Absolute abundance of genes encoding differential carbon degradation functions in plant- and microbial-derived components of different stand types"
| 组分Components | 油松纯林 P. tabuliformis forests | 辽东栎纯林 Q. wutaishanica forests | 松栎油松-辽东栎混交林 P. tabuliformis-Q. wutaishanica mixed forests | |
| 淀粉 Starch | AMY | 2.93±0.01a | 2.92±0.01ab | 2.90±0.01b |
| GBE | 2.85±0.01b | 2.90±0.01a | 2.89±0.01a | |
| GP | 3.02±0.01b | 3.11±0.02a | 3.03±0.01b | |
| 4-α-GT | 2.73±0.01ab | 2.81±0.01a | 2.74±0.01b | |
| OLG | 1.91±0.03b | 2.09±0.05a | 1.97±0.03ab | |
| 纤维素 Cellulose | β-GLU | 2.75±0.02a | 2.91±0.02ab | 2.79±0.03b |
| α-D-XYL | 2.05±0.02b | 2.14±0.02a | 2.06±0.03b | |
| 半纤维素 Hemicellulose | EX | 2.02±0.04a | 1.96±0.08ab | 1.83±0.04b |
| β-MAN | 1.88±0.02c | 2.10±0.03b | 1.99±0.02a | |
| α-GAL | 2.28±0.01b | 2.47±0.03a | 2.39±0.03a | |
| β-GAL | 1.95±0.02a | 2.11±0.06ab | 2.12±0.06b | |
| α-L-FUC2 | 2.02±0.01a | 2.03±0.04a | 1.84±0.05b | |
| 木质素 Lignin | TYR | 2.14±0.02b | 2.10±0.03b | 2.04±0.03ab |
| CAT | 2.67±0.02b | 2.82±0.04a | 2.71±0.02b | |
| CPO | 2.54±0.01c | 2.71±0.03b | 2.61±0.03a | |
| NQR | 3.14±0.01b | 3.19±0.01a | 3.14±0.01b | |
| 果胶 Pectin | α-L-RHA | 2.23±0.02a | 2.11±0.06b | 2.25±0.02a |
| 芳香族化合物 Aromatic | MIL | 1.93±0.04b | 2.05±0.02a | 2.00±0.04ab |

Fig.4
Analysis of correlations between relative abundance of CAZymes genes decomposing plant-, bacteria-, and fungi-derived substrates, and microbial biomass carbon, and environmental factors"
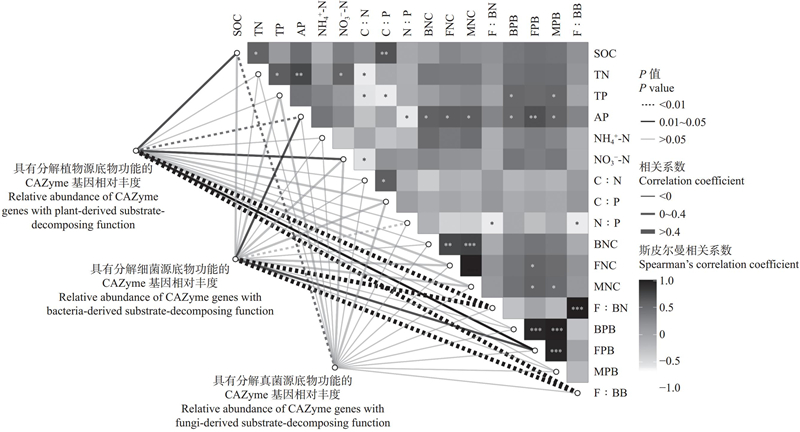

Fig.5
Association analysis of soil microbial source carbon content with relative abundance of CAZyme genes (A) and absolute abundance of carbon degradation functional genes (B)"
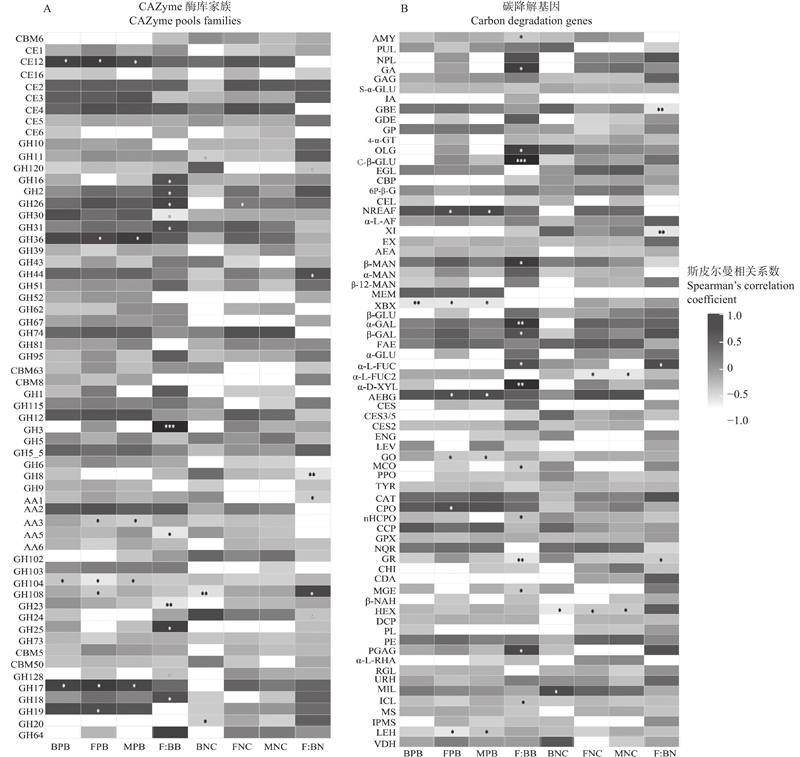

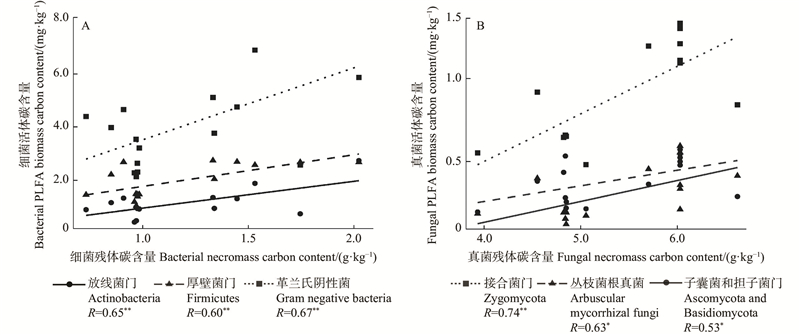
Fig.6
Regression analyses between bacterial necromass carbon content and bacterial PLFA biomass carbon content (A), and between fungal necromass carbon content and fungal PLFA biomass carbon content (B) *:P<0.05;**:P<0.01;***: P<0.001."
|
白昱欣, 刘润洪, 苏洁桦, 等. 树种混交对马尾松和红锥根际与非根际土壤微生物资源限制的影响. 生态学报, 2024, 44 (23): 10770- 10781.
doi: 10.20103/j.stxb.202310312359 |
|
|
Bai Y X, Liu R H, Su J H, et al. Effects of tree species mixing on bulk and rhizosphere soil microbial resource limitation in stands of Pinus massoniana and Castanopsis hystrix. Acta Ecologica Sinica, 2024, 44 (23): 10770- 10781.
doi: 10.20103/j.stxb.202310312359 |
|
| 淡彩虹. 2023. 黄土高原人工林混交模式对土壤团聚体稳定性及有机碳的影响. 杨凌: 西北农林科技大学. | |
| Dan C H. 2023. Effects of artificial mixed forest models on soil aggregate stability and organic carbon in the Loess Plateau. Yangling: Northwest A&F University. [in Chinese] | |
| 巩 晨. 2022. 黄土高原不同造林模式生态功能特征及权衡/协同效应. 杨凌: 中国科学院大学(中国科学院教育部水土保持与生态环境研究中心). | |
| Gong C. 2022. Characteristics and trade-off/synergy effects of ecological functions under different afforestation patterns on China’s Loess Plateau. Yangling: University of Chinese Academy of Sciences (Institute of Soil and Water Conservation Chinese Academy of Sciences and Ministry of Water Resources). [in Chinese] | |
|
樊容源, 叶绍明, 张钱春, 等. 杉木纯林及其混交林土壤团聚体活性有机碳组分特征. 西北林学院学报, 2023, 38 (6): 20- 28, 37.
doi: 10.3969/j.issn.1001-7461.2023.06.03 |
|
|
Fan R Y, Ye S M, Zhang Q C, et al. Characteristics of active organic carbon components in soil aggregates of pure plantation of C.lanceolata and its mixed forest. Journal of Northwest Forestry University, 2023, 38 (6): 20- 28, 37.
doi: 10.3969/j.issn.1001-7461.2023.06.03 |
|
| 芦 月, 杨 静, 李仕辉, 等. 2025. 黄河三角洲不同经济林土壤有机碳分布特征及改良土壤效应. 中国水土保持科学(中英文), 23(2): 180−189. | |
| Lu Y, Yang J, Li S H, et al. 2025. Distribution characteristics of soil organic carbon and soil improvement effects of different economic forests in the Yellow River Delta. Science of Soil and Water Conservation, 23(2): 180−189. [in Chinese] | |
|
王小平, 杨 雪, 杨 楠, 等. 凋落物多样性及组成对凋落物分解及土壤微生物群落的影响: 二氧化碳倍增条件下. 生态学报, 2020, 40 (17): 6171- 6178.
doi: 10.5846/stxb201812162743 |
|
|
Wang X P, Yang X, Yang N, et al. Effects of litter diversity and composition on litter decomposition characteristics and soil microbial community: under the conditions of doubling ambient atmospheric CO2 concentration. Acta Ecologica Sinica, 2020, 40 (17): 6171- 6178.
doi: 10.5846/stxb201812162743 |
|
|
严明明, 高洪娣, 刘美华, 等. 混交改造杉木纯林对林下土壤肥力影响的研究进展. 生态科学, 2024, 43 (2): 253- 259.
doi: 10.14108/j.cnki.1008-8873.2024.02.028 |
|
|
Yan M M, Gao H D, Liu M H, et al. Research progress on the effect of mixed plantation on the soil fertility in theforests between Cunninghamia lanceolata and other tree species. Ecological Science, 2024, 43 (2): 253- 259.
doi: 10.14108/j.cnki.1008-8873.2024.02.028 |
|
|
杨洪炳, 肖文发, 曾立雄, 等. 2种马尾松混交林土壤微生物特性及其与团聚体稳定性耦合关系. 江西农业大学学报, 2024, 46 (3): 543- 557.
doi: 10.3724/aauj.2024049 |
|
|
Yang H B, Xiao W F, Zeng L X, et al. Microbial characteristics in soil aggregates and their coupling with aggregates stability in two species of mixed Pinus massoniana forests. Acta Agriculturae Universitatis Jiangxiensis, 2024, 46 (3): 543- 557.
doi: 10.3724/aauj.2024049 |
|
|
姚小萌, 牛桠枫, 党珍珍, 等. 黄土高原自然植被恢复对土壤质量的影响. 地球环境学报, 2015, 6 (4): 238- 247.
doi: 10.7515/JEE201504006 |
|
|
Yao X M, Niu Y F, Dang Z Z, et al. Effects of natural vegetation restoration on soil quality on the Loess Plateau. Journal of Earth Environment, 2015, 6 (4): 238- 247.
doi: 10.7515/JEE201504006 |
|
| 于志静. 2024. 黄土丘陵区刺槐-侧柏混交林对土壤有机碳及其矿化的调控机制. 杨凌: 西北农林科技大学. | |
| Yu Z J. 2024. The regulatory mechanism of Robinia pseudoacacia-Platycladus orientalis mixed forests on soil organic carbon and its mineralization in the Loess Hilly Region. Yangling: Northwest A&F University. [in Chinese] | |
|
周建云, 李 荣, 何景峰, 等. 近自然经营对辽东栎林优势乔木更新的影响. 林业科学, 2013, 49 (8): 15- 20.
doi: 10.11707/j.1001-7488.20130803 |
|
|
Zhou J Y, Li R, He J F, et al. Regeneration of the Dominant Arbors after Close-to-Natural Management of Quercus wutaishanica Forest. Scientia Silvae Sinicae, 2013, 49 (8): 15- 20.
doi: 10.11707/j.1001-7488.20130803 |
|
|
张维伟, 尹代皓, 雷雨婷, 等. 黄土高原南部麻栎不同群落类型乔木种群结构及演替模拟. 生态学报, 2024, 44 (6): 2572- 2581.
doi: 10.20103/j.stxb.202110263011 |
|
|
Zhang W W, Yin D H, Lei Y T, et al. Structural dynamics and succession simulation of the dominant populations in three types of Quercus acutissima community in southern of the Losses Plateau. Acta Ecologica Sinica, 2024, 44 (6): 2572- 2581.
doi: 10.20103/j.stxb.202110263011 |
|
|
张兹箕, 王启鑫, 赵振宇, 等. 鲁中南地区不同经济林种植模式的改良土壤效应. 水土保持研究, 2024, 31 (5): 102- 111.
doi: 10.13869/j.cnki.rswc.2024.05.021 |
|
|
Zhang Z J, Wang Q X, Zhao Z Y, et al. Soil Improvement Effect of Different Economic Forest Planting Modes in South-central Shandong Province. Research of Soil and Water Conservation, 2024, 31 (5): 102- 111.
doi: 10.13869/j.cnki.rswc.2024.05.021 |
|
|
Azeem M, Sun T R, Jeyasundar P G S A, et al. Biochar-derived dissolved organic matter (BDOM) and its influence on soil microbial community composition, function, and activity: a review. Critical Reviews in Environmental Science and Technology, 2023, 53 (21): 1912- 1934.
doi: 10.1080/10643389.2023.2190333 |
|
|
Bourget M Y, Fanin N, Fromin N, et al. Plant litter chemistry drives long-lasting changes in the catabolic capacities of soil microbial communities. Functional Ecology, 2023, 37 (7): 2014- 2028.
doi: 10.1111/1365-2435.14379 |
|
|
Cudjoe E, Ruiz-Peinado R, Pretzsch H, et al. Neighborhood competition improves biomass estimation for Scots pine (Pinus sylvestris L.) but not Pyrenean oak (Quercus pyrenaica Willd. ) in young mixed forest stands. Forest Ecosystems, 2025, 13, 100317..
doi: 10.1016/j.fecs.2025.100317 |
|
|
Choreño-Parra E M, Treseder K K. Mycorrhizal fungi modify decomposition: a meta-analysis. New Phytologist, 2024, 242 (6): 2763- 2774.
doi: 10.1111/nph.19748 |
|
|
Duarte A G, Maherali H. A meta-analysis of the effects of climate change on the mutualism between plants and arbuscular mycorrhizal fungi. Ecology and Evolution, 2022, 12 (1): e8518.
doi: 10.1002/ece3.8518 |
|
|
Hu D, Xu Y, Chai Y F, et al. Spatial Distribution pattern and genetic diversity of Quercus wutaishanica Mayr population in Loess Plateau of China. Forests, 2022, 13 (9): 1375.
doi: 10.3390/f13091375 |
|
|
Fan X Y, Ge A H, Qi S S, et al. Root exudates and microbial metabolites: signals and nutrients in plant-microbe interactions. Science China Life Sciences, 2025, 68, 2290- 2302.
doi: 10.1007/s11427-024-2876-0 |
|
|
Fu X H, Song Q L, Li S Q, et al. Dynamic changes in bacterial community structure are associated with distinct priming effect patterns. Soil Biology and Biochemistry, 2022, 169, 108671.
doi: 10.1016/j.soilbio.2022.108671 |
|
|
Gedminienė L, Spiridonov A, Stančikaitė M, et al. Temporal and spatial climate changes in the mid-Baltic region in the Late Glacial and the Holocene: Pollen-based reconstructions. Catena, 2025, 252, 108851.
doi: 10.1016/j.catena.2025.108851 |
|
|
Gao G, Huang X M, Xu H C, et al. Conversion of pure Chinese fir plantation to multi-layered mixed plantation enhances the soil aggregate stability by regulating microbial communities in subtropical China. Forest Ecosystems, 2022, 9, 100078.
doi: 10.1016/j.fecs.2022.100078 |
|
|
Han Z G, Zhou Y C, Guo Y L, et al. Organic carbon sequestration by Fe oxides in aggregates in a Pinus massoniana conifer-broadleaf mixed forest. Soil and Tillage Research, 2024, 240, 106105.
doi: 10.1016/j.still.2024.106105 |
|
|
Hu D, Zhou X H, Ma G Y, et al. Increased soil bacteria-fungus interactions promote soil nutrient availability, plant growth, and coexistence. Science of The Total Environment, 2024, 955, 176919.
doi: 10.1016/j.scitotenv.2024.176919 |
|
|
Huffman M S, Madritch M D. Soil microbial response following wildfires in thermic oak-pine forests. Biology and Fertility of Soils, 2018, 54, 985- 997.
doi: 10.1007/s00374-018-1322-5 |
|
|
Jiang F Y, Zhang L, Zhou J C, et al. Arbuscular mycorrhizal fungi enhance mineralisation of organic phosphorus by carrying bacteria along their extraradical hyphae. New Phytologist, 2021, 230, 304- 315.
doi: 10.1111/nph.17081 |
|
|
Joergensen R G. Amino sugars as specific indices for fungal and bacterial residues in soil. Biology and Fertility of Soils, 2018, 54 (5): 559- 568.
doi: 10.1007/s00374-018-1288-3 |
|
|
Kakouridis A, Yuan M T, Nuccio E E, et al. Arbuscular mycorrhiza convey significant plant carbon to a diverse hyphosphere microbial food web and mineral-associated organic matter. New Phytologist, 2024, 242, 1661- 1675.
doi: 10.1111/nph.19560 |
|
|
Kohler J, Roldán A, Campoy M, et al. Unraveling the role of hyphal networks from arbuscular mycorrhizal fungi in aggregate stabilization of semiarid soils with different textures and carbonate contents. Plant and Soil, 2017, 410, 273- 281.
doi: 10.1007/s11104-016-3001-3 |
|
|
Liang C, Amelung W, Lehmann J, et al. Quantitative assessment of microbial necromass contribution to soil organic matter. Global Change Biology, 2019, 25 (11): 3578- 3590.
doi: 10.1111/gcb.14781 |
|
|
Mercer G D, Mickan B S, Gleeson D B, et al. Probing the pump: soil carbon dynamics, microbial carbon use efficiency and community composition in response to stoichiometrically-balanced compost and biochar. Soil Biology and Biochemistry, 2025, 205, 109770..
doi: 10.1016/j.soilbio.2025.109770 |
|
|
Manici L M, Caputo F, Fornasier F, et al. Ascomycota and Basidiomycota fungal phyla as indicators of land use efficiency for soil organic carbon accrual with woody plantations. Ecological Indicators, 2024, 160, 111796.
doi: 10.1016/j.ecolind.2024.111796 |
|
|
Mou Z J, Kuang L H, Zhang J, et al. Nutrient availability and stoichiometry mediate microbial effects on soil carbon sequestration in tropical forests. Soil Biology and Biochemistry, 2023, 186, 109186.
doi: 10.1016/j.soilbio.2023.109186 |
|
|
Starke R, Mondéjar R L, Human Z R, et al. Niche differentiation of bacteria and fungi in carbon and nitrogen cycling of different habitats in a temperate coniferous forest: a metaproteomic approach. Soil Biology and Biochemistry, 2021, 155, 108170.
doi: 10.1016/j.soilbio.2021.108170 |
|
|
Tao F, Huang Y Y, Hungate B A, et al. Microbial carbon use efficiency promotes global soil carbon storage. Nature, 2023, 618, 981- 985.
doi: 10.1038/s41586-023-06042-3 |
|
|
Toriyama J, Hashimoto S, Nakao K, et al. Management strategies for shrinking and aging tree plantations are constrained by the synergies and trade-offs between carbon sequestration and other forest ecosystem services. Journal of Environmental Management, 2025, 373, 123762.
doi: 10.1016/j.jenvman.2024.123762 |
|
|
Wang B R, Huang Y M, Li N, et al. Initial soil formation by biocrusts: Nitrogen demand and clay protection control microbial necromass accrual and recycling. Soil Biology and Biochemistry, 2022, 167, 108607.
doi: 10.1016/j.soilbio.2022.108607 |
|
|
Wiesenbauer J, Gorka S, Jenab K, et al. Preferential use of organic acids over sugars by soil microbes in simulated root exudation. Soil Biology and Biochemistry, 2025, 203, 109738.
doi: 10.1016/j.soilbio.2025.109738 |
|
|
Zhang L M, Wu Y N, Yan L B, et al. Modeling the maximal organic carbon saturation capacity in karst forest soils of China: comparing the existing models. Applied Ecology and Environmental Research, 2021, 19 (5): 4115- 4128.
doi: 10.15666/aeer/1905_41154128 |
| [1] | Xiao Li,Shuxian Jia,Yingqing Xi,Liuming Yang,Xiaofei Liu. Effects of Litter Addition and Removal on Soil Microbial Necromass Carbon in a Natural Forest of Castanopsis carlesii [J]. Scientia Silvae Sinicae, 2024, 60(10): 12-20. |
| [2] | Zi Hongbiao, Xiang Zeyu, Wang Genxu, Ade Luji, Wang Changting. Profile of Soil Microbial Community under Different Stand Types in Qinghai Province [J]. Scientia Silvae Sinicae, 2017, 53(3): 21-32. |
| Viewed | ||||||
|
Full text |
|
|||||
|
Abstract |
|
|||||